1. Symmetry and Size
Annelids are all bilaterally symmetrical animals. They range in size from much less than 1 mm in length to more than 3 m. The smallest annelids described to date would appear to be Neotenotrocha (Dorvilleidae, Eunicida) reaching only a few hundred microns in length (Eibye-Jacobsen and Kristensen, 1994). Also within Eunicida may be found the largest of annelids. A preserved specimen of Eunice (Eunicidae) at the Museum of Natural History in Washington D. C. is close to 3 m in length and around 5 cm in diameter in its preserved and contracted condition. Some earthworms, such as the Giant Gippsland Earthworm (Megascolides australis), can reach 1 m in length. The largest leech in the world, Haementeria ghilianii, is found in South America and reaches more than 40 cm in length.
2. Coelom
Nearly all annelids have a fluid-filled cavity between the outer body wall and the gut, and this is referred to as a coelom (Figure 1). The coelom is often used as a storage area for gametes and acts as a hydrostatic skeleton for locomotion. The coelom (formed via schizocoely) is generally organized as a series of compartments divided by intersegmental septa. This compartmentalization means that if the worm is damaged the coelomic contents will only be lost from a few segments, and locomotion can be maintained. In other annelids there may only be a few septa dividing the coelom. Under these circumstances much more coelomic fluid is lost with injury, and the worm may be severely affected. A number of groups, particularly leeches and a number of small polychaetes, have little or no coelomic space.

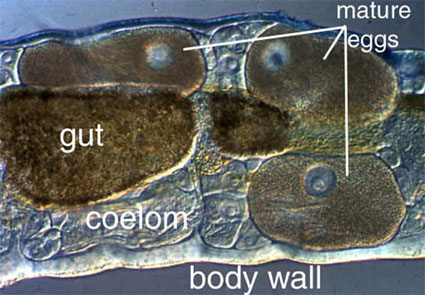
Figure 1. Amphicorina mobilis (Sabellidae). Bondi, Australia. Copyright © 1996 Greg Rouse.
3. Body wall
Annelids have a body covered by an external cuticle that is never shed or molted. Epidermal microvilli secrete a network of fibers that are in part collagenous and also contain scleroprotein. Chaetae are also cuticular structures, but contain large amounts of chitin. Beneath the epidermis and its basal lamina lies a layer of circular muscle. The circular muscle layer forms a nearly continuous sheath around the body, except in polychaetes with well-developed parapodia. Beneath the circular muscle layer lie thick longitudinal muscles. In many annelids the longitudinal muscles are present as four distinct bands (Fig. 2).
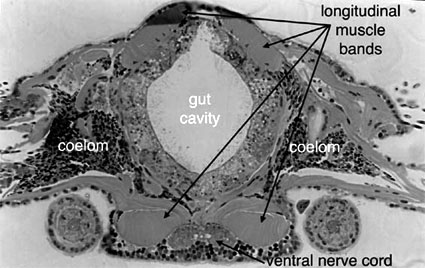
Figure 2. capricornia (Hesionidae) (Pleijel and Rouse, 2000). One Tree island, Australia. Copyright © 2000 Greg Rouse.
4. Parapodia
Parapodia are unjointed segmental extensions of the body wall found in many polychaetes, though many do lack these features. They are also absent in Clitellata and Echiura. Parapodia are equipped with musculature derived mainly from the circular muscle layer and usually carry chaetae. Parapodia vary in structure but basically can be considered to consist of two elements; a dorsal notopodium and a ventral neuropodium (Fig. 3). In addition to bundles of chaetae, noto- and neuropodia can also have a variety of cirri and gills. They are most elaborate in actively crawling or swimming forms where they form large fleshy lobes that act as paddles. Parapodia of burrowing or tubicolous polychaetes can simply be slightly raised ridges carrying hooked chaetae called uncini.
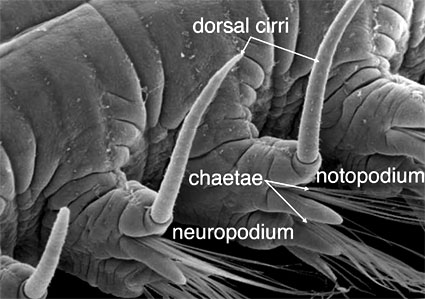
Figure 3. Ophiodromus flexuosus (Hesionidae). Banyuls, France. Copyright © Fredrik Pleijel.
5. Nervous system
Annelids have a brain or cerebral ganglion that originates and usually resides in the head. The brain varies in structure, with mobile active forms having the most complex brains, and sessile or burrowing forms having simple brains with little differentiation. It is connected to the ventral nerve cord by the circumpharyngeal connectives, which run down each side of the pharynx. The ventral nerve cord, is usually made up of a pair of cords that are bound together and runs the length of the body (Fig. 4). It varies in thickness and dilates into a ganglion in each segment, from which pairs of segmental nerves pass out to the body wall, muscles and gut.
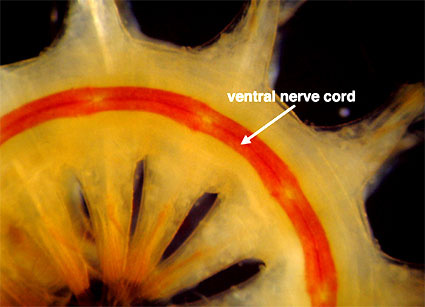
Figure 4. Ventral view of a scaleworm (Sigalionidae) showing its double nerve cord. This worm is unusual in having red pigmentation associated with the cord, making it easily seen. Misaki, Japan. Copyright © 1998 Greg Rouse.
6. Sense organs
There are six major kinds of sensory structures found in annelids. These include palps, antennae, eyes, statocysts, nuchal organs and lateral organs (Fig. 5). Palps and antennae are located on the head of many polychaetes. In some groups they are both sensory while in others the palps are used for feeding. Nuchal organs are ciliated, paired, chemosensory structures, innervated from the posterior part of the brain. They are present in nearly all polychaetes, and Rouse and Fauchald (1997) suggested that they may represent an apomorphy for Polychaeta. This has been challenged by other authors who suggest that nuchal organs may be an apomorphy for Annelida as a whole and have been lost in Clitellata/Oligochaeta (Purschke et al., 2000). Annelids also have a variety of epidermal sensory cells that may be responsive to light or touch (such as lateral organs).

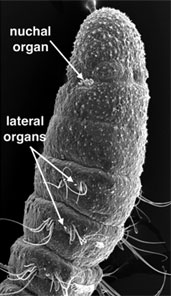
Figure 5. Left, Eumida (Phyllodocidae). Bundeena, Australia. Copyright © 1999 Greg Rouse. Right, Proscoloplos (Orbiniidae). Bondi, Australia. Copyright © 2001 Greg Rouse.
7. Circulation and respiratory structures
A closed circulatory system is present in most polychaetes (Fig. 6), Echiura and many clitellates. It is reduced or absent in leeches where it may be replaced by coelomic canals. A limited circulatory system in which some of the major blood vessels are present but the distal capillary vessels are missing is found in a number of polychaete groups. A circulatory system is absent in many small polychaetes.
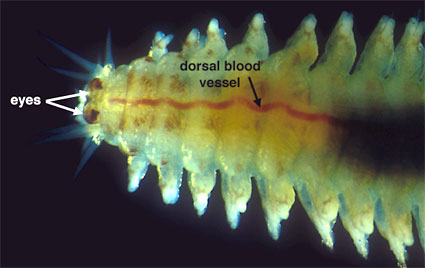
Figure 6. Australonereis ehlersi (Nereididae). Bundeena, Australia. Copyright © 1999 Greg Rouse.
8. Segmental organs
In most annelids there are usually two fluid systems, the coelom and the circulatory system, and both (if present) are involved in the excretion of waste products. To achieve this excretion there must be ducts to the exterior, and these are generally referred to as nephridia. Ducts, known as gonoducts or coelomoducts, are also required for the transfer of gametes that develop in the coelom to the outside of the body on maturity. The two different kinds of ducts are often simply referred to as segmental organs since determining what kind of duct is present is problematic.